









Μ. Δ. Υφαντίδης
Προοίμιο
Τον Ιανουάριο του τρέχοντος έτους δημοσιεύτηκε (σε ηλεκτρονική μορφή) στα Proceedings of the National Academy of Sciences των ΗΠΑ διδακτορική διατριβή με τίτλο “Varroa destructor feeds primarily on honey bee fat body tissue not hemolymph”9 (“Η Varroa destructor τρέφεται πρωτίστως με λιπώδη ιστό της μέλισσας, όχι με αιμολέμφο”). Ωστόσο, στα τέλη του 2018 ο Samuel. Ramsey κατά την καθιερωμένη, προφορική υποστήριξη αυτής της διατριβής του ενώπιον ακροατηρίου στο πανεπιστήμιο του Maryland την είχε τιτλοφορήσει με τον αυθεντικό και κάπως πιο «ανατρεπτικό» τίτλο:
«Varroa does not feed on hemolymph»
,ο οποίος σε σχετικό βίντεο του διαδικτύου προβάλλεται φαρδύς-πλατύς στην γιγαντο-οθόνη της κεντρικής αίθουσας διαλέξεων του ως άνω πανεπιστημίου.
Έχουμε λοιπόν καταρχήν δημοσιοποίηση μιας και της αυτής ερευνητικής εργασίας με δύο όμως αμφίσημους τίτλους, κάτι που είναι απότοκο της χρήσης κειμένου ηλεκτρονικής μορφής. Για όσο ακόμη χρονικό διάστημα θα παραμένει το κείμενο μόνο με τη μορφή αυτή, θα υπάρχει και η δυνατότητα της όποιας άλλης (και κατά το δοκούν) τροποποίησής του. Πράγματι, στη δεύτερη, μετριοπαθέστερη εκδοχή, της υπό σχολιασμό εργασίας9 προστέθηκε και η ακόλουθη διατύπωση-ομολογία των δώδεκα συνολικά συν-συγγραφέων: «Δεν είμαστε βέβαιοι ότι η βαρρόα δεν καταναλίσκει και αιμολέμφο».
Με τον ένα ή τον άλλον τίτλο, αυτή η επιστημονική είδηση για το είδος τροφής της βαρρόα προκάλεσε, είναι η αλήθεια, το εύλογο ενδιαφέρον του αναγνωστικού κοινού, παγκοσμίως. Μόνο μέσα στην πρώτη εβδομάδα του Φεβρουαρίου του 2019 ο αριθμός των επισκεπτών της σχετικής ιστοσελίδας αυξήθηκε κατά 536 άτομα, φτάνοντας έτσι έως τότε συνολικά στους 15.958 αναγνώστες. Εκλαϊκευτικά μελισσοκομικά περιοδικά πρόβαλαν ήδη και σχολίασαν ποικιλοτρόπως την περίεργη, όντως, αυτή είδηση. Βρισκόμαστε άραγε στην αρχή της δημιουργίας ενός καινούριου μελισσοκομικού μύθου, ογδόντα περίπου χρόνια μετά την εποχή, κατά την οποία γεννήθηκε εκείνος, ο περίφημος πια (αλλά τουλάχιστον εξαιρετικά θελκτικός ! ) μύθος της γλώσσας των χορών των μελισσών;
Πάντως, το συμπέρασμα-αφορισμός του Ramsey, ότι δηλαδή «η βαρρόα δεν τρέφεται με αιμολέμφο», (ο αυθεντικός τίτλος της εργασίας του) δικαιολογημένα ξαφνιάζει, διότι μοιάζει σαν να ανατρέπει άρδην την επιστημονικά καθολική παραδοχή, ότι δηλαδή το παράσιτο τρέφεται με την υγρής μορφής αιμολέμφο (με το «αίμα») της μέλισσας. Η ανακάλυψη ότι η τροφή της ακμαίας βαρρόα είναι πρωτίστως ο μη υγρής μορφής λιπώδης ιστός της (επίσης) ακμαίας μέλισσας συνιστά αναμφίβολα πρωτοτυπία της διδακτορικής αυτής διατριβής, και διευρύνει, ασφαλώς, όμως χωρίς και να ανατρέπει τις ως τώρα γνώσεις μας πάνω στο συγκεκριμένο θέμα. Η καινούρια αυτή γνώση πιθανολογείται, ότι θα συμβάλει θετικότατα στην παραπέρα κατανόηση της συμπεριφοράς της βαρρόα κατά τη διαδικασία της μετακίνησής της από το σώμα της ακμαίας μέλισσας στα ώριμα κελιά του ανοιχτού γόνου του μελισσιού, μέσα στα οποία και αναπαράγεται αποκλειστικά το παράσιτο αυτό. Ωστόσο, η ανακάλυψη του Ramsey ατυχώς δεν προσδίδει αυτομάτως στην εν λόγω διατριβή και το αναμενόμενο επιστημονικό κύρος, διότι, το αρχικό συμπέρασμα ότι η βαρρόα δεν τρέφεται με αιμολέμφο», είναι τελείως αβάσιμο.
Τα τρωτά σημεία της διατριβής του Ramsey
1.Η απολυτότητα στη διάκριση αιμολέμφου και λιπώδη ιστού
Τα ως σήμερα βιβλιογραφικά δεδομένα πείθουν, ότι αιμολέμφος και λιπώδης ιστός είναι δύο «συγκοινωνούντες» ιστοί (οργανικά συστήματα) της μέλισσας, και ότι ανάλογα με το στάδιο της οντογενετικής εξέλιξης του εντόμου τροφοδοτεί ο ένας ιστός τον άλλον με θρεπτικά συστατικά 1,2,11,14. Πιο συγκεκριμένα, στο στάδιο της ακμαίας μέλισσας η αιμολέμφος της (που περιλούζει όλους τους άλλους ιστούς και τα όργανα του εντόμου, άρα και τον λιπώδη ιστό) εμπλουτίζεται πρωτογενώς με θρεπτικά συστατικά, προερχόμενα κατ’ ευθείαν από την τροφή (μέλι και γύρη) με την αφομοιωτική δράση των κυττάρων του μονόστρωμου επιθηλίου του στομάχου του εντόμου. Στη συνέχεια η ίδια η αιμολέμφος εφοδιάζει αδιάλειπτα με αυτά τα συστατικά τους υπόλοιπους ιστούς του. Ότι περισσεύει, αποθηκεύεται στον λιπώδη ιστό. Αντίθετα, στο στάδιο της αναπτυσσόμενης μέλισσας μέσα στο σφραγισμένο πλέον κελί του γόνου, και ειδικότερα στο στάδιο της προχρυσαλίδας, είναι ο ήδη σχηματισμένος πλούσιος λιπώδης ιστός της που εφοδιάζει με τη σειρά του την αιμολέμφο με αφθονία καλοχωνεμένων ουσιών. Τότε είναι που ο λιπώδης ιστός, καθώς ωριμάζει, εξαντλείται πλήρως, δηλαδή, «μετατρέπεται» σε αιμολέμφο, όπως παραστατικά δείχνει και η Εικόνα 1: τα λιποσταγονίδια [λευκοί κύκλοι στα στάδια α), β), και γ)] λιγοστεύουν υπέρ των υδατανθρακούχων και πρωτεϊνούχων ουσιών [σκούροι κύκλοι στα στάδια δ) και ε) ] 11.

Από τα προαναφερθέντα γίνεται τώρα, μάλλον, κατανοητό ότι καταρχήν μεταξύ αιμολέμφου και λιπώδη ιστού υπάρχει μια δυναμική ισορροπία και όχι, (όπως αντίθετα πρεσβεύουν οι συγγραφείς της υπό ανάλυση εδώ εργασίας) μια στατική διαφορά. Ως προς την χημική σύσταση των δύο αυτών ιστών η διαφορά τους δεν είναι τόσο ποιοτική όσο ποσοτική. Και οι δυο, δηλαδή, περιέχουν υδατάνθρακες, πεπτίδια (πρωτεϊνες) και λιπαρές ουσίες, αλλά σε διαφορετικές αναλογίες. Ως προς τη φυσική τους κατάσταση, εξάλλου, η διαφορά των δύο ιστών έγκειται στο ότι μόνον η αιμολέμφος είναι ρευστή και γι αυτό μπορεί και κυκλοφορεί μέσα στο σώμα του εντόμου, ενώ ο μη υγρός λιπώδης ιστός αυτονόητα στερείται της σπουδαίας αυτής φυσιολογικής ιδιότητας. Να γιατί, λοιπόν, το δίλημμα «αιμολέμφος ή λιπώδης ιστός» ως τροφή της βαρρόα καταρχήν είναι πλαστό.

Πάντως, ο λιπώδης ιστός της ακμαίας μέλισσας (Εικόνα 2), τον οποίο κατά κύριο λόγο καταναλίσκει η ακμαία βαρρόα9 αποτελείται από ελαφρά συνδεμένες μεταξύ τους, λευκές λιποκύστεις (lipocytes), οι οποίες έχουν και αυτόνομη συνθετική δραστηριότητα η καθεμιά τους. Πράγματι, στα έντομα γενικά, αλλά και στη μέλισσα ειδικότερα, ο λιπώδης ιστός δεν είναι απλώς αποθήκη “λίπους” και άλλων οργανικών και ανόργανων ουσιών, αλλά και όργανο, το οποίο διαδραματίζει ανάλογο ρόλο με το συκώτι των θηλαστικών, δηλαδή, σε αυτόν λαμβάνει χώρα η σύνθεση (με τη διαδικασία του λεγόμενου ενδιάμεσου μεταβολισμού) όχι μόνο λιπών, με τη μορφή λιποσταγονιδίων, αλλά επίσης και γλυκογόνου και πρωτεϊνών. Επιπλέον, ο λιπώδης ιστός είναι και ενδοκρινικό όργανο, το οποίο παράγει διάφορα αντιμικροβιακά πεπτίδια και συμμετέχει στην αποτοξίνωση του οργανισμού του εντόμου, για παράδειγμα, από υπολείμματα εντομοκτόνων, κ.λπ.1
- Ανεπαρκής βιβλιογραφία στο θέμα «τροφή των αναπτυσσόμενων βαρρόα»
Αυτό είναι το δεύτερο τρωτό σημείο της εργασίας του Ramsey και των συνεργατών. Όπως επισημάνθηκε ήδη και νωρίτερα, ο λιπώδης ιστός «ωριμάζει», κατά το διάστημα που η προχρυσαλίδα της μέλισσας μεταμορφώνεται6 σε χρυσαλίδα (βλ. και πάλι Εικόνα 1). Τότε τα ώριμα λιποκύτταρα βαθμιαία διογκώνονται αρχικά λόγω του οργανικού νερού, που παράγεται κατά τον ενδιάμεσο μεταβολισμό των λιπαρών ουσιών, και τελικά διαρρηγνύονται αναπότρεπτα, οπότε το περιεχόμενό τους εμπλουτίζει αυτομάτως την αιμολέμφο11. Σε αυτή τη φάση της οντογενετικής εξέλιξης της μέλισσας, ο λιπώδης ιστός της έχει πράγματι μετατραπεί φυσιολογικά ολόκληρος σε αιμολέμφο. Αυτό επισημαίνεται και από την ερευνητική ομάδα στην δεύτερη εκδοχή, της υπό ανάλυση εργασίας, ότι δηλαδή «Ο λιπώδης ιστός εξαντλείται για τις ανάγκες δημιουργίας των νέων οργάνων της χρυσαλίδας»9 (Ramsey και συνεργάτες 2019, σελ..4, στήλη 2, παρ. 2). Η «εξάντληση» του λιπώδη ιστού συμβαίνει προφανώς υπό εντελώς φυσιολογικές συνθήκες (είτε, δηλαδή, η μέλισσα παρασιτείται από τη βαρρόα είτε όχι). Σε κάθε περίπτωση, αυτή η «εξάντληση» εξηγεί, γιατί ο λιπώδης ιστός των νέο-εκκολαπτόμενων μελισσών είναι όντως υποτυπώδης έως και ανύπαρκτος, όπως απερίφραστα δηλώνεται και πάλι από τους συγγραφείς της υπό ανάλυση διδακτορικής διατριβής (βλ. σελ 4, στήλη 2 παράγραφος 1 σειρές 8-10: «… νεοεκκολαπτόμενες μέλισσες έχουν εξαντλημένο τον λιπώδη ιστό (λόγω των αναγκών της μεταμόρφωσης…)» Τον πιο καλά αναπτυγμένο λιπώδη ιστό, πάντως, τον διαθέτουν οι οικιακές μέλισσες, ηλικίας άνω της μιας εβδομάδος (Εικόνα 2). Σε αυτές τις μέλισσες είναι που παρασιτεί και ο μεγαλύτερος αριθμός των ακμαίων ακάρεων9.
Επισημαίνεται τέλος ότι οι αναπτυσσόμενες βαρρόα εμφανίζονται στο σφραγισμένο κελί της μέλισσας ως πρωτονύμφες, (το πρώτο κινητικό οντογενετικό στάδιο του παρασίτου 8,12) και άρα αυτές τότε αρχίζουν για πρώτη φορά να διατρέφονται, όταν, δηλαδή, η αναπτυσσόμενη μέλισσα θα έχει ήδη μεταμορφωθεί σε χρυσαλίδα, με άλλα λόγια τότε που θα έχει «εξαντληθεί» ήδη ο λιπώδης ιστός και όχι νωρίτερα. Αντιλαμβανόμαστε λοιπόν, ότι οι αναπτυσσόμενες βαρρόα τρέφονται ΜΟΝΟΝ με αιμολέμφο, αφού τότε δεν υπάρχει λιπώδης ιστός στις αναπτυσσόμενες μέλισσες Επομένως ο νεότευκτος μύθος του Ramsey, δηλαδή, ότι «Η βαρρόα δεν τρέφεται με αιμολέμφο» καταπίπτει αυτομάτως εν τη γενέσει του.
Πώς παρασιτούν οι αναπτυσσόμενες βαρρόα
Πάντως, το υπό ανάλυση ζήτημα αποκτά την ουσιαστικότερη σημασία του, όταν αναλογιστούμε ότι την εξάντληση του μελισσιού, ως υπεροργανισμού, δεν την προκαλεί ο παρασιτισμός από τις ακμαίες αλλά κυρίως από τις αναπτυσσόμενες βαρρόα, όπως θα φανεί κατά την πρόοδο της συζήτησής μας.
Είναι, λοιπόν, κατ’ αρχήν διαπιστωμένο, ότι η ικανότητα του παρασίτου να τραυματίζει με τα προς τούτο κατάλληλα διαμορφωμένα στοματικά του μόρια5 τον ξενιστή του είναι «προνόμιο» μόνον της ακμαίας βαρρόα. Τα στοματικά μόρια των αναπτυσσόμενων βαρρόα είναι κατάλληλα μόνον για αναρρόφηση υγρής τροφής (αιμολέμφου). Πάντως, τα τραύματα τα προκαλεί το παράσιτο στο πιο ευάλωτο σημείο της ακμαίας μέλισσας (εργάτριας ή κηφήνα), δηλαδή, στις λεπτές, μεμβράνες, που συνδέουν τους κοιλιακούς δακτυλίους του εντόμου μεταξύ τους. Οι αναπτυσσόμενες βαρρόα, σε αντίθεση με τη μητέρα τους, όχι μόνον δεν μπορούν να τραυματίσουν τον ξενιστή τους (δηλαδή, την αναπτυσσόμενη τώρα μέλισσα), αλλά στερούνται και του μηχανισμού της λεγόμενης έξω-στοματικής πέψης (extra-oral digestion)3. Με τον μηχανισμό αυτό, και πάλι μόνον η ακμαία βαρρόα πολτοποιεί τα κύτταρα του λιπώδη ιστού (όπου τον βρει) και στη συνέχεια καταπίνει τον βλωμό. Ο πολύ απλής δομής πεπτικός αγωγός της βαρρόα δεν διαθέτει αδενικά κύτταρα, που να παράγουν λιπολυτικά ένζυμα3.
Εύλογα, λοιπόν θα μπορούσε να αναρωτηθεί τώρα κανείς, πώς μπορούν και διατρέφονται τα εξελισσόμενα ακάρεα μέσα στο κλειστό ακόμα κελί της αναπτυσσόμενης μέλισσας, ενώ δεν μπορούν καν να ανοίξουν πληγές στο σώμα του ξενιστή τους και ενώ δεν διαθέτουν ούτε ένζυμα, που να μπορούν να μετουσιώνουν τον λιπώδη ιστό σε αιμολέμφο;. Η απάντηση στο ερώτημα αυτό έχει δοθεί εδώ και μερικές δεκαετίες κατά σαφή και αδιαμφισβήτητο τρόπο με την πρωτοποριακή και εξαιρετικής σημασίας εργασία των Donze & Querin (1994)4, την οποία, ωστόσο, δεν μνημονεύουν διόλου οι Ramsey και συνεργάτες (2019). Η γονική βαρρόα, δηλαδή, ενώ μέσα στο σφραγισμένο κελί του γόνου καταναλίσκει τον άγουρο ακόμη λιπώδη ιστό της προχρυσαλίδας11 (Εικ.3α) τρυπώντας την σε διάφορα, τυχαία σημεία του σώματός της, περιορίζει τη λήψη της τροφής και για την ίδια αλλά και για τους απογόνους της σε ένα μόνον σημείο του σώματος του ξενιστή, όταν αυτός βρίσκεται πλέον στο στάδιο της χρυσαλίδας11 (Εικόνα 3β), όταν, δηλαδή, ο λιπώδης ιστός είναι ώριμος, και έχει πλέον μετουσιωθεί σε αιμολέμφο, όπως ήδη έχει τονιστεί. Εκεί η ακμαία βαρρόα δημιουργεί μια και μόνη πληγή, μια μικροσκοπική «γούρνα» στο σώμα της χρυσαλίδας, όπου μπορούν να γευματίζουν και οι αναπτυσσόμενες βαρρόα.
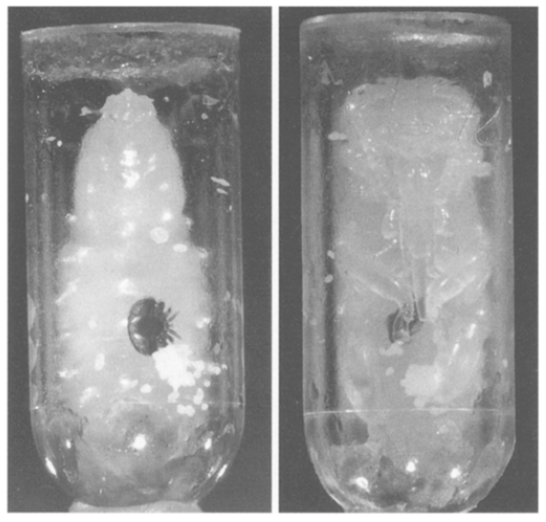
Το φαινόμενο, που σύντομα περιγράφηκε εδώ, έχει χαρακτηριστεί ως η «γονική μέριμνα» της βαρρόα4. Το τυχόν ad hoc επιχείρημα (με τέτοια συντηρούνται όλοι οι μύθοι !) προς υποστήριξη και του νεότευκτου μύθου του Ramsey, ότι δηλαδή η γονική βαρρόα φτάνει τάχα στο σημείο να «θηλάζει» τα μωρά της με πολτοποιημένο από την ίδια λιπώδη ιστό είναι ανίσχυρο, καθώς μάλιστα υπάρχει και ανταγωνισμός στη θέση της λήψης της τροφής, όχι μόνον μεταξύ των απογόνων αλλά και ανάμεσα σε αυτούς και στην μητέρα4. Αλλά, φαινόμενα «θηλασμού» έτσι κι αλλιώς δεν παρατήρησαν οι Donze & Querin (1994)4
Αποτελεί πάντως για τη βαρρόα, ως ζωολογικό είδος, ένα εξαιρετικά σημαντικό, επιλεκτικό πλεονέκτημα το γεγονός, ότι την ανοιχτή πληγή την δημιουργεί το παράσιτο στο υψηλότερο σημείο του (σε ύπτια θέση) σώματος της χρυσαλίδας της μέλισσας, δηλαδή, στο έπαρμα του πέμπτου σωματικού δακτυλίου (Εικόνα 4).
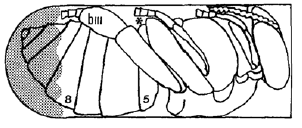
Εξ αυτού του λόγου διασφαλίζεται το γεγονός ότι ο ξενιστής (παρόλον ότι η αιμολέμφος δεν πήζει 1, 2, 11, 14) «δεν διατρέχει τον κίνδυνο να πεθαίνει από ακατάσχετη αιμοραγία», όπως πολύ εύστοχα έχει ήδη διατυπωθεί και από προγενέστερους συγγραφείς4. Αυτός ο κίνδυνος για τον ξενιστή (συνεπώς εμμέσως πλην σαφώς και για το ίδιο το παράσιτο) μειώνεται πιθανότατα και εξ αιτίας της διαδικασίας της ψευδο-επούλωσης της πληγής με τη συνάθροιση εκεί αιματοκυστών της αιμολέμφου 2. Η πληγή κλείνει πλήρως, όταν η χρυσαλίδα μεταμορφώνεται σε ακμαία μέλισσα με τον σχηματισμό του νέου εξωσκελετού της.
Η πρακτική πλευρά του ζητήματος
Η ζημία που υφίσταται πρωτογενώς ένα μελίσσι από τη βαρρόα προκαλείται πράγματι από τα αναπτυσσόμενα ακάρεα. Δευτερογενώς η βαρρόα μπορεί να καταστρέφει το μελίσσι και με την διάδοση ιώσεών του. Πάντως, σε ατομικό επίπεδο αναγόμενη η απώλεια σε αιμολέμφο της παρασιτούμενης μέλισσας είναι ασύγκριτα μικρότερη, όταν προκαλείται από μια ακμαία παρά από μια αναπτυσσόμενη βαρρόα. Και τούτο διότι η αφαίμαξη της ακμαίας μέλισσας συντηρεί απλώς το ακμαίο παράσιτο στη ζωή, ενώ η αφαίμαξη της αναπτυσσόμενης μέλισσας συνεπάγεται και την παραγωγή βιομάζας [με τη μορφή των βαρρόα της καινούριας γενεάς]. Πράγματι, βρέθηκε ότι οι νεοεκκολαπτόμενες, παρασιτούμενες μέλισσες ζυγίζουν σημαντικά λιγότερο από εκείνες, που αναπτύσσονται σε κελιά χωρίς βαρρόα ή σε κελιά που είναι προσβεβλημένα από άγονες βαρρόα10. Αυτή η σημαντική απώλεια αιμολέμφου της αναπτυσσόμενης μέλισσας είναι όντως και η ρίζα του κακού, η κύρια αιτία, δηλαδή, για το ότι ένα προσβεβλημένο από βαρρόα μελίσσι των ευρωπαϊκών φυλών της Apis mellifera καταρρέει τελικά μέσα σε τρία χρόνια, αν μείνει αβοήθητο από το μελισσοκόμο.
Από την ως τώρα συζήτηση προκύπτει επίσης ότι οι αναπτυσσόμενες βαρρόα καταναλώνοντας αποκλειστικά και μόνον αιμολέμφο εξαντλούν σε πολύ σοβαρό βαθμό την αναπτυσσόμενη μέλισσα (χρυσαλίδα), μάλιστα και μέχρι θανάτου (σε περιπτώσεις πολλαπλών προσβολών του ίδιου κελιού γόνου από γόνιμες βαρρόα). Από πρακτική, λοιπόν, άποψη σημασία έχει να αναπτύξουμε μεθόδους καταπολέμησης της βαρρόα, με τις οποίες να προκαλέσουμε τη μεγαλύτερη δυνατή νεανική θνησιμότητα του παρασίτου. Οι μέθοδες αυτές δεν χρειάζεται να είναι χημικές, όπως αντίθετα διατείνονται οι συγγραφείς της αναλυθείσας τώρα διατριβής, οι οποίοι και ευαγγελίζονται τη δημιουργία διασυστηματικών ακαρεοκτόνων, που θα ενσωματώνονται στον λιπώδη ιστό της ακμαίας μέλισσας. Αυτήν την επιδίωξή τους μάλιστα την τονίζουν ιδιαίτερα στην εισαγωγή της εργασίας τους : «Ο στόχος μας, παρέχει (διανοίγει)το μονοπάτι προς ανάπτυξη νέων στρατηγικών θεραπείας για τη βαρρόα» (σελ. 1, στήλη 2, οι τελευταίες δύο σειρές της τελευταίας παραγράφου)9. Χάριν αυτού του στόχου όμως το επιστημονικό κύρος της συγκεκριμένης ερευνητικής εργασίας έχει τραυματιστεί ανεπανόρθωτα. Από την άλλη μεριά, όπως απέδειξε η μελισσοκομική πράξη δεκαετιών, η φιλοσοφία της χημικής καταπολέμησης του παρασίτου είναι μάλλον ατελέσφορη και αδιέξοδη, αφού η «βαρρόα» παραμένει ακόμη η νούμερο 1 απειλή της μελισσοκομίας, σε μελίσσια των Ευρωπαϊκών φυλών της A. mellifera7. Αναμφίβολα, όμως, η χημειοθεραπεία είναι ευεργετική οικονομικά, πρώτον για τις ίδιες τις φαρμακοβιομηχανίες, που χρηματοδοτούν αδρά τις σχετικές έρευνες για ανακάλυψη νέων φαρμάκων, δεύτερον για τους αναλυτές μελιού, που έχουν έτσι δια βίου εξασφαλισμένη εργασία για τον αρκετά μάλιστα δαπανηρό προσδιορισμό καταλοίπων, και τρίτον για τους ίδιους τους μελισσοκόμους, αφού και για αυτούς είναι πολύ προσφιλείς οι διαδικασίες, που ανταποκρίνονται στο μοτίβο: «ψεκάστε, σκουπίστε, τελειώσατε», ή αλλιώς στο μοτίβο «ο χρόνος είναι χρήμα».
Πάντως, το γενικότερο συμπέρασμά μας είναι, ότι οι ως τώρα δημοσιεύσεις για την φυσιολογία της θρέψης της μέλισσας 1,2,6,14, αλλά και οι σχετικές ερευνητικές εργασίες και βιβλία για την αναπαραγωγή και τη διατροφή της βαρρόα 8, 12,13 δεν χρειάζεται να ξαναγραφούν, όπως αντίθετα με έκδηλη μάλιστα την σκωπτική διάθεση τους ισχυρίζονται οι Ramsey και συνεργάτες (2019)9.
Βιβλιογραφία
1,. Arrese, E.L., Soulages,J. L. 2011. Insect fat body: Energy, Metabolism, and Regulation. HHS Public Access.
2., Chapman, R. F. 1999. The Insect structure and function. The English University Press LTD, 5th Edition London.
- Cohen, A. C. 1955. Extra-oral digestion in predaceous terrestrial Arthropoda. Annual Review of Entomology, 40:85-103
- Donze, G., Querin, M.P.,1994. Behavioral attributes and parental care of Varroa mites parasitizing honeybee brood. Behavioral and Ecological Sociobiology, 34: 305-319.
- Griffiths D.A.1988. Functional morphology of the mouthparts of Varroa jacobsoni Africanized Honeybees and Bee Mites pp 479-486.
- Jenkin, P. M. 1970. Control of growth and metamorphosis. Pergamon Press, Oxford.
- Locke, B. 2016. Natural Varroa mite-surviving Apis mellifera honeybee populations.Apidologie 47:467-482.
- Martin, S.J. 1994. Ontogenesis of the mite Varroa jacobsoni Oud. In worker brood of the honey bee Apis mellifera L. under natural conditions. Experimental and Applied Acarology, 18(2):87-100.
9.Ramsey, S.D., Ochoa, R., Bauchan, G., Gulbonson, C., Mowery J.D, Cohen, A., Lim, D., Joklik J., Cicero, J.M., Ellis, J.D., Hawthorne, D., Engelsdrop von D., 2019. Varroa destructor feeds primarily on honey bee fat body tissue not hemolymph. Proceedings of the National Academy of Sciences of U.S.A.
10.Schneider, P., Drescher, W. 1987. Einfluss der Parasitierung durch die Milbe Varroa jacobsoni Oud. Auf das Schlupfgewicht, die Gewichtsentwicklung, die Entwicklung der hzpopharynxdrüsen und die Lebensdauer von Apis mellifera L. Apidologie, 18(1)¨101-110.
11.Seifert , R. W. 1975. Entomologisches Praktikum. Georg Thieme-Verlag Stuttgart
12.Υφαντίδης, Μ Δ. 1983. Ontogenesis of the mite Varroa jacobsoni Oud. In worker and drone honeybee brood cells. Journal of Apicultural research, 22(3):227-233.
13, Υφαντίδης, Μ.Δ. 2011.Παθήσεις του μελισσιού-Μη συμβατικές μέθοδοι αντιμετώπισής τους. Εκδόσεις Ν. Παππάς σελ. 333
- Wigglesworth, V.B. 1995. The Principles of Insect Physiology, London:Methuen & Co, LTD..
![]()
Απορίες και διευκρινίσεις σχετικά με το άρθρο ή με όποιο άλλο θέμα θέλετε επικοινωνήστε με το περιοδικό Μελισσοκομική Επιθεώρηση πατώντας εδώ.
Για την κατάρτισή σας,
επιλέξτε πηγές πληροφόρησης με επιστημονικά τεκμηριωμένη γνώση